


The brain as a statistician
Sensory coding 1: phototransduction


As we saw last week, animals’ ability to turn sensory information into behavior defines many of the computational problems we’d like to solve. How would one design a neural system to perform these computations? We’ll explore sensory coding with two examples: phototransduction in the eye (this post), and acoustic transduction in the ear (next post).
In every environment, we are constantly bombarded by electromagnetic waves. Anthropocentrically, we call waves at 400-700nm light1. But how do we convert light into electrical signals and glutamate release that eventually drives perception and action? Phototransduction in rods, which are photoreceptors in the retina used in low luminance conditions, illustrates how the brain is able to discriminate between relevant signals — photons from the world — and noise — spontaneous activation in a rod.
Although we think of light as waves, under low luminance, the retina senses light intensity by counting the number of photons. In moonlight, rods catch photons at a rate of about one per second. In darker environments, this rate has been psychophysically determined to be about one every 5000 seconds per rod in humans. To accomplish this, a rod must very reliably detect a single photon!
Inside a rod, rhodopsin molecules catch photons, isomerize (change chemical configuration), and initialize a biochemical cascade. Rhodopsin must be sensitive enough to detect one photon, but because strong collisions between molecules can also cause isomerization, they must be stable enough as they float around in the cell and bump into each other. When a collision does set off the isomerization, it appears to downstream processes identical to a photon catch. This is potentially bad!
Do these collisions happen often enough for us to care? For one rhodopsin molecule, the rate is about once every 700 years. Phew, you might say. BUT! There are hundreds of millions of rhodopsin molecules per rod, so this rate turns out to be about once every 160 seconds for a rod. Even worse, taking into account all the rods in the retina means that these events happen quite frequently. This noise is called spontaneous isomerization, often perceived as a scintillating pattern. And of course the Germans have a word for it: eigengrau, or intrinsic gray. A simulation of the eigengrau on a 300x300 patch of rods:

Here’s the remarkable thing: the one photon per 5000 seconds per rod threshold was determined from the conditions under which a subject can discriminate between actual light and no light with 75% accuracy. How can the threshold be this low if spontaneous isomerizations happen at a much higher rate of once every 160 seconds per rod? How is it possible that a tiny change in rate is enough for the brain to say: this is light!
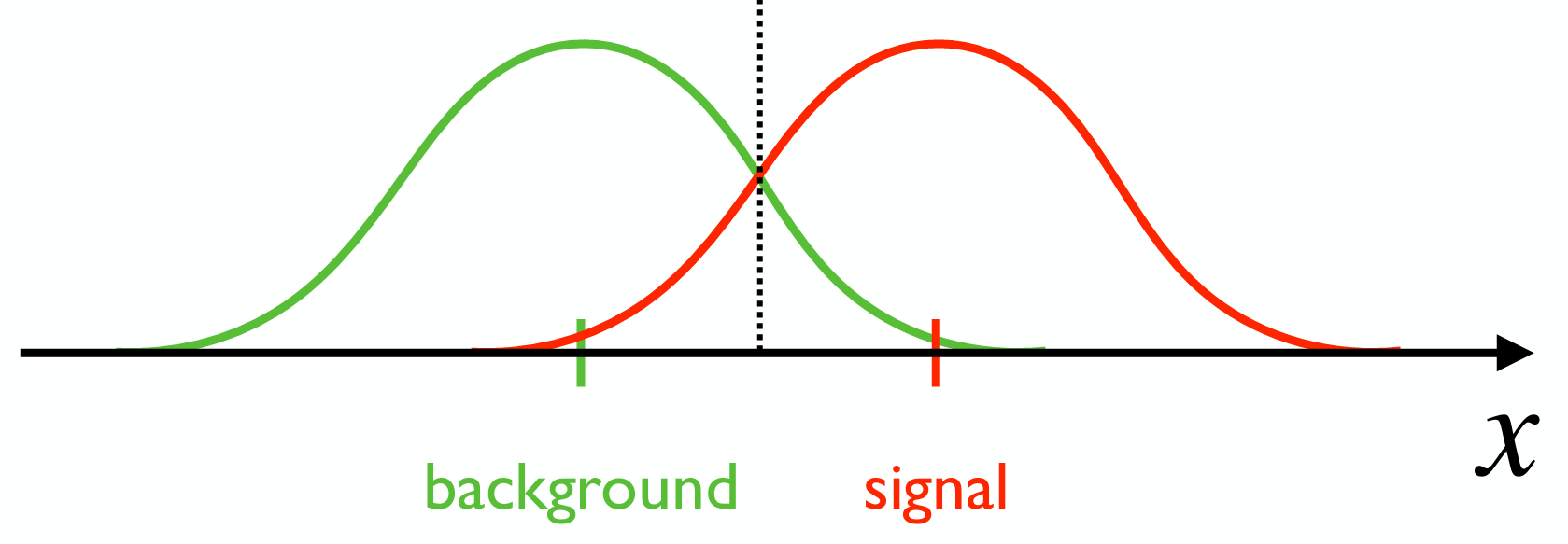
Let’s express this slightly more formally. We can model the background and signal distributions as Poisson, with λ_b=1/160 as the background spontaneous isomerization rate and λ_s=(1/160 + 1/5000) as the signal rate. Given an event, the brain must determine which distribution caused it. If the two distributions are very separated like in the cartoon below, maybe this is not too hard. You could be correct a high percentage of the time by using the dotted line below as a threshold. Say you perceived x amount of brightness: if it was to the left of the line, you say there’s no real light. Otherwise, you say there’s light. Even if you move the line, there would be some tradeoff between true and false positives, but you would not be that wrong2.
In reality, the two rates only differ by about 3%. Even at a much higher rate of λ_b=10 (where λ_s also increases to maintain the 3% difference), the distributions are very close. At λ_b=100, they are a little bit farther. For both settings, using a threshold line to classify between the two distributions would result in a very high chance of being wrong.

Because the signal and background always differ by 3%, the goal is to increase the absolute distance by raising the rate such that the 3% is enough to get high accuracy by drawing a line. Qualitatively, it should look something like this:

Note that we cannot literally increase the 1/5000 threshold rate; this is a fixed property of the visual system. However, we could integrate over some number of rods and some period of time, i.e. count the number of events that occur in a spatiotemporal region, until we get an aggregated rate that achieves the desired accuracy. I will not give exact numbers in case future versions of this course use the same problem set, but it is a ton of rods and a surprisingly long amount of time3! Amazingly, the brain seems to have solved this problem: it is able to do these statistics constantly and as correctly as necessary.
This balance of sensitivity and stability continues to be important downstream of the rods. Bipolar cells in the retina must reliably detect a photon amongst a pool of rods. If just one rod catches a photon, there must somehow be a reliable change in the voltage of the bipolar cell. The challenge is, as Bruno says, to constantly fight the entropy of the universe, such as random fluctuations in synaptic terminals. If you were to linearly sum the noisy rod responses within each pool together, an individual photon event wouldn’t be detectable. The solution is to place thresholds at synapses, which results in passing only a real light signal (as much as possible) to the bipolar cell. What is the optimal threshold? Work by Sampath, Field, and Rieke4 calculated a theoretical optimal value based on the amount of noise at each synaptic terminal and the signal magnitude in response to actual light. Experimentally, they confirmed that the threshold was exactly where theory said it would be!
Perhaps this all seems like a convoluted, nonsensical system. But in the spirit of building a brain, the challenge would be: could you come up with a better mechanism that operates with such low energy, with such a high dynamic range, under these constraints? Really good cameras don’t have to work like the eye, but it’s still important (and fun, hopefully) to understand the principles underlying how biology solved this problem.
In the next post, we will see a solution to another problem: acoustic transduction.
A bee would include the ultraviolet portion of the spectrum; a mosquito or snake, infrared.
I didn’t want to go into area under the ROC curve (bold black curve below) in detail, but at a high level, the 75% accuracy refers to the area under the curve generated by every location of the threshold line for the two distributions. Each threshold line has a different tradeoff between true positives (hits) and false positives, which each fall somewhere on the ROC curve. If the area under the curve is 0.75 (out of 1), then the distributions are appropriately far apart. As an example, assuming the distributions are far enough apart, the threshold line below corresponds to the dot on the curve.

This is part of why rods do not sense fast temporal changes as well as cones do.
Nonlinear signal transfer from mouse rods to bipolar cells and implications for visual sensitivity (Field & Rieke 2002); Selective Transmission of Single Photon Responses by Saturation at the Rod-to-Rod Bipolar Synapse (Sampath & Rieke 2004).